
|
|
|
Research Article
Quantifying the response of tree hyraxes (Dendrohyrax validus) to human disturbance in the Udzungwa Mountains, TanzaniaJ. Elmer Topp-Jørgensen1, Andrew R. Marshall2,3,* , Henry Brink4 and Ulrik B. Pedersen1 1Danish Zoological Society, Denmark, e-mail: [email protected]. 2Centre for Ecology Law and Policy, Environment Department, University of York, Heslington, York YO10 5DD, UK, e-mail: [email protected]. 3Flamingo Land, Kirby Misperton, Malton, North Yorkshire YO17 6UX, UK. 4Durrell Institute of Conservation and Ecology, University of Kent, Canterbury, Kent CT2 7NS, UK, e-mail: [email protected]. *Author for correspondence.
Received: 16 January 2008, Accepted: 1 February 2008, Published: 3 March 2008 Copyright: This is an open access paper. We use the Creative Commons Attribution 3.0 license http://creativecommons.org Cite this paper as: Topp-Jørgensen, J. E., Marshall, A. R.., Brink, H. and Pedersen, U. B. 2008. Quantifying the response of tree hyraxes (Dendrohyrax validus) to human disturbance in the Udzungwa Mountains, Tanzania. Tropical Conservation Science 1 (1):63-74. Available online: tropicalconservationscience.org Introduction The number of species threatened with extinction in tropical forests is predicted to increase [1]. Most forms of disturbance are undetectable or only marginally detectable using remote methods [2]. Assessing the consequences of disturbance by ground surveys is therefore a major priority for conservation management. Fully quantifying the effects of disturbance would require painstaking work due to the massive number of species involved. However, basic criteria for assessing ecosystem health and habitat composition and structure are rarely determined [3]. Selection of key species as "indicators," "guilds," or "functional types" can assist in making more rapid assessments [4-7]. Determining the impact of disturbance on rare species is also of importance in determining habitat requirements for management.
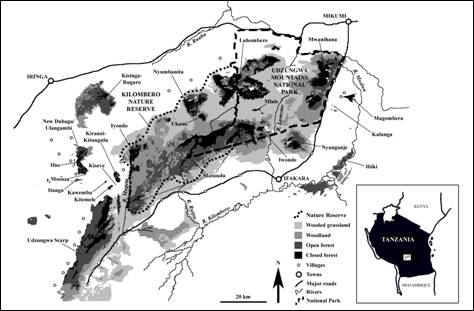
There have only been three published field surveys of the ecology of D. validus, including one in the Eastern Arc Mountains. There have also been no attempts to make direct counts of D. validus density nor to determine how disturbance affects behavior. Furthermore, there have been no studies to compare methods for estimating density. This lack of research is regrettable because the hyrax's uneven density and patchy distribution has to be understood before practical conservation plans can be developed. A common approach has been to estimate hyrax latrine density [11,12]. One such study, in two of the same areas as the current study, showed that the latrine density of hyrax in the West Kilombero Scarp Forest Reserve was far greater than in the heavily hunted New Dabaga / Ulangambi Forest Reserve. A second study in South Africa also showed that den preference of D. arboreus is closely linked to diet [13]. From both of these studies it is clear that tall, intact forest is crucial for D. validus survival. The study aim is to determine the sensitivity of D. validus to human disturbance. If D. validus is a forest-dependent species, we would expect a negative relationship with disturbance. Here we compare relative density and behavior from three reserves in one of the world's most important areas for biodiversity. From this we assess the impact of destructive human activity both within and between sites. We also evaluate three methods for estimating D. validus relative density; namely circular plots, transect counts, and indirect counts using latrines. Methods Study Sites The Udzungwa Mountains contain the largest area of forest of all the Eastern Arc blocks (1,500 km2). Like most tropical forests, a large area has been degraded by humans (62%; [14, 15]; Fig. 2). Three Forest Reserves differing in hunting level and logging history were selected for comparison of hyrax density. The reserves are located in the same general habitat (montane and upper montane forest) and exposed to similar rainfall (1500-2000 mm yr-1) and temperature regimes (10-27°C; [16]. Tree species overlap is high (>50 species between the three reserves [17] including several genera—e.g. Pododcarpus, Teclea, Hagenia, Ficus, Strychnos, Apodytes, Rhus—that are known to be used for shelter by the southern tree hyrax Dendrohyrax arboreus). [10, 13, 18].
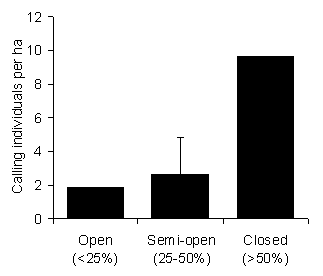
At the time of survey, the first of our study sites was known as the West Kilombero Scarp Forest Reserve (WKSFR). However, this was later upgraded to Nature Reserve status (the highest protective status under Tanzanian Forestry and Beekeeping Division legislation). The new Kilombero Nature Reserve comprises WKSFR and two other former Forest Reserves (Iyondo and Matundu) [19], including several forest fragments (Fig. 1). The reserve has a high elevation range; however, our study site was at 1825 m. KNR contains fragmented forest that is among the least disturbed by humans in the Udzungwa Mountains, and previous expeditions have recorded a high diversity of large mammals [17,20,21]. Although small-scale poaching occurs in the reserve, this is mostly restricted to grassland, woodland, and forest edge areas. Hunters target larger game, but also smaller species, including hyrax (personal communication with hunters). Levels of hunting are lower than the other two study sites. 2. Udzungwa Scarp Forest Reserve (USFR) The second study area, in the north of USFR at 1900 m elevation, was in an area known as Kihanga. The area comprises 230 km2 of forest, of which around 100 km2 is closed-canopy [14]. Two well-used footpaths run through the survey area, from Mbawe and Masisiwe villages on the plateau, to the Kilombero Valley. During 36 field days, 31 people were counted on the main paths, dogs accompanying people were recorded twice, and gunshots were heard on two occasions. Signs of hunting, observed along 4.26 km of transect and approximately 11 km of paths, included nine pitfalls (two active), 19 snares (five active), two huts containing snare string, and 15 trees cut to force D. validus to the ground (1.88 ha-1; Topp-Jørgensen and Pedersen unpublished data). 3. New Dabaga/ Ulangambi Forest Reserve (NDUFR) NDUFR is a 37.28 km2 reserve with 32.0 km2 of forest (12.8 km2 closed-canopy) [14] on an undulating plateau near the village of Kidabaga at 1900 m elevation. It has a long history of commercial exploitation of timber, which has altered the structure and tree species composition of the forest (18.45 pitsaw sites/ha; [22]). Villages and agricultural land surrounded NDUFR long before its gazettement in the 1930s. Numerous east-west paths cross the reserve between villages. Hence many people traverse the forest regularly and hunting is common throughout. An average of 32.6 active traps km-2 (snares and pitfalls) were recorded 5 m either side of 40.5 km of transect [22]. However, no trees felled for catching hyrax were observed. Interviews with hunters revealed an annual harvest of 45.5 hyrax per km2 [12]. Relative Density We used the circular plot count (or "point count") method [23,24]. Loud calls made within 50 m of a fixed point were recorded within the first hour of calling after dusk. The Udzungwa Mountains are a particularly difficult area for estimating density due to undulating terrain and poor visibility due to tangled climbers [25]. To account for this, the fixed points were placed away from steep slopes or ridges and also away from noisy waterways. Sample sites were also restricted to areas easily accessible from suitable campsites (< 500 m), to avoid walking long distances in the dark. Campsite locations were selected to maximize the spread of samples. Finally, points were positioned at least 100 m from campsites to avoid human interference, in forest with the most intact canopy available. The survey was initiated from the first call heard within the census radius. For each call, the time, compass direction, and a distance estimate were recorded. To minimize error in distance estimation, all observers practiced estimating distances against distances measured using a tape measure. Observers also spent time familiarizing themselves with the forest structure and location/height of trees likely to contain hyrax before each count. This allowed observers to consider the likelihood of a call being outside of the 50 m radius, e.g., by considering the angle of the slope to the call, versus the location of known trees. At the end of every count, the minimum number of calling individuals within the 50 m radius was calculated as the number of calls that certainly came from separate individuals. This minimum estimate was largely based on common sense, using the spatial and temporal proximity of calls. Wherever hyrax were present in or near plots, counts were repeated at least once (up to four repetitions), to verify the estimate. Counts were conducted by Topp-Jørgensen and Pedersen at five sites in KNR (Ndundulu) and four sites in USFR. These counts were made between the short and long rainy seasons (November 1997 to March 1998). In 2000, six circular plot counts were also made in NDUFR by Topp-Jørgensen, Marshall, and Brink, between July and October. Despite this being the "dry" season, regular showers occurred every week. Hyrax latrines were also counted in all three reserves, to evaluate their use as a measure of hyrax relative density. A latrine was defined as any pile of hyrax dung containing both fresh and dry pellets. Latrines were recorded within 5 m of transects giving a 10 m strip. 4.26 ha were censused in KNR, 4.10 ha in USFR, and 4.65 ha in NDUFR. Finally, hyrax calls and sun-basking individuals were recorded to estimate relative density and to detect possible behavioral changes in daytime activity induced by hunting. We made 22 daytime (07:00-11:30 h) repeated walks along a 4.42 km transect in undisturbed KNR and 21 walks along a 4.26 km transect in USFR, walking at a pace of 1 km h-1. All observations of hyrax were recorded, noting whether visual or vocal. When comparing forests, we use the term "relative density" instead of "true density", to emphasize that the number of individuals is unknown. For circular plot and transect counts, this would require knowledge of the proportion of calling-to-non-calling individuals. For latrine counts the proportion of hyrax defecating in latrines versus those defecating randomly would be required. Furthermore, the number of individuals sharing latrines would be needed. Canopy Structure Human disturbance can have a major impact on canopy structure. This can occur by direct removal of stems, secondary damage caused by fallen stems, and increased elephant activity encouraged by rapid regeneration of herbaceous vegetation. Assessment of the relationship between canopy structure and hyrax was carried out at 9 sites with varying canopy structure in areas of KNR at least 4 km from settlements (5 in Ndundulu, 3 in Nyumbanitu and 1 in Ukami). KNR was chosen to minimize bias from hunting, hence using only sites that were a long distance from settlements. Three observers (Topp-Jørgensen, Marshall, and Brink) carried out plot count surveys in the dry season between July and November 2000. Three repetitions were made at each site and mean density of calling individuals was calculated for use in all analyses. Canopy structure was assigned to one of three categories at every site. Structure categories were defined by canopy cover estimated by eye (i.e., the proportion of sky obscured by branches of tall trees within a 50 m radius). We use these broad categories rather than a continuous scale to minimize bias from estimating cover by eye (Fig. 3): 1) Open canopy (<25% canopy cover): These sites had few mature trees, dense vines entangling regenerating trees, and few arboreal pathways between trees—2 plots. 2) Semi-open canopy (25-50% canopy cover): These sites had a mix of regenerating and mature trees, dense shrubby vegetation, and some arboreal pathways between canopies—5 plots. 3) Closed canopy (>50% canopy cover) - these plots included mature, closed canopy forest with numerous arboreal pathways—2 plots.
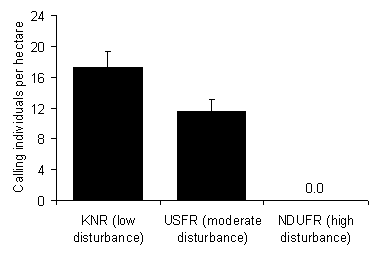
Study sites were spread across the elevation gradient (1140 m to 2100 m elevation), at varying distances from the forest edge. We also therefore tested the relationship between the density of calling individuals and these two factors, to determine whether they may have had any influence on the observed trends. Kolmogorov-Smirnoff tests revealed that all datasets were not significantly different from a normal distribution and thus parametric tests are used throughout. Results Relative Density The number of calling individuals in circular plots had a positive relationship with disturbance (mean 17.3 ± 1.9 ha-1 95 % confidence interval in the little disturbed KNR, 11.7 ± 1.4 ha-1 in moderately disturbed USFR, and 0.0 ± 0.0 ha-1 in the heavily hunted and formerly logged NDUFR; Fig. 4; Table 1). Although none were heard during plot counts, calls were heard outside of plots on rare occasions in NDUFR. Density was significantly higher in KNR than USFR (t-test: t=4.43, df=7 p=0.003). As no hyraxes were heard calling in NDUFR plots, density was not estimated. No significant difference in call rate per individual was observed between KNR wet season (Feb-March 1998) and the dry season canopy structure selection data (July-August 2000; t-test: t=1.149, df=10, p=0.277). Furthermore, calls were heard every night from July through March in KNR, a period containing both wet and dry seasons. This suggests that the seasonal difference in surveys does not explain the difference in density estimates between KNR, USFR, and NDUFR. Table 1. Density of hyrax calls and latrines and visual/vocal observations per transect walked (mean ± 95 % confidence interval), from three forests in the Udzungwa Mountains. Also given is the relative density (% in parentheses) to that seen in KNR, for each method.
Fixed-area searches revealed 11 and zero latrines in KNR and USFR, respectively (2.6 and 0.0 ha-1; Table 1). The relative density of USFR compared to KNR is far less for latrines than for calling individuals (0.0 % versus 67.6 %; Table 1). One latrine was, however, found casually near to the census area in USFR. No latrines were observed in NDUFR. Hyrax visual and vocal observations made during daytime transect walks were too few to estimate density. In KNR, D. validus was seen at 10 different positions (45.5 % of walks) and calls were heard on 21 out of 22 walks (95.5 %; Table 1). In USFR, no hyraxes were seen during the 21 transect walks and calls were only heard during three of these (14.3 %). Therefore, as for latrine counts, daytime transect walks in USFR suggest a relative density to KNR far less than for circular plots (0.0 % visual and 15.0 % vocal, versus 67.6 %; Table 1). They also suggest that D. validus engages less in sun-basking and daytime calling in USFR compared to the less hunted KNR.
Discussion We assume that the observed differences between forests are due to human influences. The site with the lowest relative density (NDUFR) was the most degraded and hunted. The observed relative density between NDUFR and KNR confirms previous findings for these two forests [12]. The site with intermediate relative density (USFR) also had intermediate disturbance. The three methods used agree broadly with these relative differences. However the density estimate and magnitude of the differences vary widely between methods, and only circular plot counts of calls could clearly differentiate the densities of USFR and NDUFR. This has implications for method selection, which we now discuss along with the implications of disturbance for D. validus. Disturbance and Relative Density The results agree with a previous study, that hunting is a major concern for mammals in these areas [12]. However, we feel that habitat is also a major consideration. Given that hunting and habitat degradation tend to occur together, these two forms of disturbance are often hard to separate [33]. However, we have shown here that D. validus in KNR is more abundant in undisturbed closed canopy forest than in more open forest. Hunting levels were not thought to vary between these same sites as all were distant from human settlements, with extensive areas of more accessible forest in between. Studies suggest that tree hyrax density is limited by cavity-bearing trees used for shelter [13,18]. Preferred shelter trees are large, part of the upper canopy, partly decayed, have several entrance holes, and are usually of dietary value [13]. Home-range size for D. arboreus has been shown to increase with a decrease in tree density [18]. Tree hyraxes frequently spend time on the ground [18], but also depend on arboreal pathways [8]. The lack of arboreal pathways in open forest may increase vulnerability to hunting by snares or terrestrial predators. Avoidance of open-forest structure types may therefore reflect both habitat unsuitability and an anti-hunting/predation strategy. However in NDUFR, D. validus was also absent from many areas with intact, mature forest cover, suggesting that other factors may be involved (e.g., hunting or disease). Besides human disturbance, inter-specific competition with the yellow-spotted bush hyrax (Heterohyrax brucei) may have contributed to the reduced density of D. validus in NDUFR. However, the presence of this species was not known for KNR and USFR. The bush hyrax is not a typical moist-forest species, but logging may have allowed this widespread species, more typical of drier habitats [27], to enter NDUFR. Disturbance and Behaviour Given that there were many calling individuals in USFR, the lack of defecation sites in this forest suggests that latrines have either been reduced or abandoned. This could be a behavioral response to hunting. In NDUFR, logging may also have removed traditional defecation sites. In a Uganda study of D. arboreus, 43 of 127 latrines were inactive [18], further highlighting the uncertainty of this method. Three earlier studies of tree hyraxes reported them to be nocturnal [8,30,31], but later studies have also revealed examples of diurnality [32] and equal activity level day and night [26]. In the undisturbed KNR, D. validus was frequently heard calling and observed basking on branches during the day. However no animals were observed during the daytime in the hunted forests (USFR and NDUFR) and few calls were heard during daytime. Hunting therefore seems to shape the activity pattern, making it more difficult for the hunter to locate the animal during the day. The circular plot data is also interesting for annual variation in behavior. Seasonality in nocturnal calling behavior has been found for both D. validus [10] and D. arboreus [26]. In the Virunga Massif (Uganda), calls were heard on 52 out of 61 nights during the dry season and on 17 out of 100 nights in the wet season [18]. However there was no evidence of a birth season. The lack of obvious seasonality for D. validus in high elevation Udzungwa (this study), further suggests that seasonality in calling may be shaped by environmental and climatic conditions. Comparison to Previous Studies Densities of hyraxes recorded elsewhere suggest that the density of calling individuals found in our circular plots (17.3 ± 1.9 ha-1 KNR; 11.7 ± 1.4 ha-1 USFR) may be feasible estimates of true density, as they fall within previously published density ranges. Our densities of latrines (2.6 ha-1 KNR; 0.0 ha-1 USFR) are, however, very low compared to the literature. Mark-recapture of D. arboreus in Rwanda found 10.3 adults and 3.8 juveniles ha-1 in undisturbed montane forest [26]. However, latrine counts of D. validus on Kilimanjaro found between eight and 70 ha-1 [11]. The extremely high upper estimate of the Kilimanjaro study was made in an area with 153 trees above 32 cm dbh per hectare. D. arboreus in South Africa prefer shelter trees > 35 cm dbh [13]. If true for D. validus, this would mean that hyraxes occurred in 45.8 % of trees in the Kilimanjaro study. This seems high considering that hyraxes depend on cavity-bearing trees and avoid some species (including Juniperus procera and Cassipourea malosana, which made up 14.4 % of trees in the Kilimanjaro study). In addition to the observed variation in relative density estimates within our study, the density of latrines seen for KNR also differs from that reported by the only previous study in this area [12]. From transect counts, 6.06 ± 2.92 latrines ha-1 were found in KNR [12], as compared to our 2.6 ha-1. The reason for this difference is unknown. The range of forest habitat types in KNR is high and the length of transect surveyed by the earlier study in KNR was greater (19.40 km versus 4.26 km). The difference may therefore be due to sample size, location, and/or the definition of a latrine (the definition used by the earlier study is not given [12]). However, like our survey, no latrines were found in NDUFR.
Table 2. Comparison of non-invasive methods for estimating density of hyraxes.
In addition to our major aim of determining hyrax responses to disturbance, the results also provide a guide to methods for counting hyraxes. Non-invasive methods for estimating relative density of hyraxes are compared in Table 2. From the methods tested, we prefer plot counts of calling individuals at dusk, as dusk calling behavior seems to vary less than latrine-use and daytime calling. Daytime transect counts are impractical for tree hyraxes as they are seen and heard too infrequently during daylight hours. Night transect walks are a potential alternative; however, these may be dangerous in areas with difficult terrain or a high density of large animals, such as KNR. The drawback of circular plots is gaining accurate measurements. A major assumption of distance sampling (i.e., determining densities through estimation of distances) for both transect and point counts, is that distances are measured accurately [34-36]. However, the distance from the central point of our circular plots was estimated only from the sound of calling. While many efforts were made to minimize bias, there is still potential for error. This uncertainty in the circular plot method could be improved by having additional observers (or microphones), in a triangle or square shape to better pinpoint the location of calls. Using microphones could further help to discern individuals. Night-vision or heat-sensing equipment would also improve estimates from any plot or transect based surveys made after dark. To summarize, in the absence of true density estimation we have shown that relative density can be a suitable alternative, as is sometimes the case for other mammals, e.g., primates [25]. However, further work is needed to compare relative density estimation methods, preferably versus known density (e.g. through mark-recapture). Ongoing efforts to explore methods for other mammals could be used as a model [28,29,33]. Implications for Conservation It has been suggested that selective logging could help finance tree hyrax conservation by logging only tree species not used for shelter or food [13]. However in light of the D. validus relationship with canopy cover, the role of forest structure should be investigated more closely. This is particularly important in areas with significant ground trapping, where the resulting forest structure change (regardless of the species logged), may reduce hyrax density and thus counteract conservation efforts. Furthermore, bushmeat is of low importance for human nutrition in the area [12]. The dangerously low density in NDUFR also shows the unsustainability of current hunting practices. Therefore hunting is an inappropriate management strategy. More importantly than the conservation of a single species, the study also has implications for conservation of biodiversity-rich forest habitats. Typically, forest reserves are not considered part of the global protected area network [37]. Here we have demonstrated the major reason why, i.e., forest reserves are protected by legislation but not by active management. As a result illegal activities such as hunting and pole-cutting can continue unabated and unquantified. The effect seen here for hyraxes has been mirrored by several other taxa in the Udzungwa Mountains [12,14,17,29]. The creation of Kilombero Nature Reserve [19] and initiation of joint forest management, since conducting our surveys, opens the future possibility for monitoring the effect of management intervention. Acknowledgments The Åge V. Jensen Foundation sponsored Topp-Jørgensen and Pedersen in KNR and USFR. We also thank the University of Dar es Salaam, Tanzania Wildlife Research Institute, and Tanzanian Commission for Science and Technology. Thanks to Helle Jørgensbye Hansen, Martin R. Nielsen, and Prof. J. Fjeldså for providing comments on early drafts. In the field, we were guided and helped by many villagers, especially Elia Mulungu, Janus Mdanga, Mzee Mohando, and Lactali Mehwa. Thanks also to Dave Switzer, Mark Burrage, and Dave Naish for valuable input during fieldwork. In addition to data compiled by the authors for KNR and USFR, this publication uses material collected during the Udzungwa Mountains Biodiversity Surveys; a collaboration between the Society for Environmental Exploration and the University of Dar es Salaam (Frontier Tanzania), and the Udzungwa Mountains Forest Management and Biodiversity Conservation Component, MEMA, supported by the Danish Government through DANIDA. References [1] Whitmore, T.C. and Sayer, J.A. 1992. Tropical Deforestation and Species Extinction. Chapman and Hall, London. [2] Peres, C.A., Barlow, J. and Laurance, W.F. 2006. Detecting anthropogenic disturbance in tropical forests. Trends in Ecology and Evolution 21:227-229. [3] Balmford, A., Green, R. E. and Jenkins, M. 2003. Measuring the changing state of nature. Trends in Ecology and Evolution 18:326-330. [4] Skorupa, J.P. 1986. Responses of rainforest primates to selective logging in Kibale Forest, Uganda. In: K. Benirschke (ed.) Primates: The Road to Self-Sustaining Populations. Springer-Verlag, New York, 57-70. [5] Skorupa, J.P. 1988. The effects of Selective Timber Harvesting on Rain-forest Primates in Kibale Forest, Uganda. Ph.D. Thesis, University of California, Davis, USA. [6] Landres, P.B., Verner, J. and Thomas, J.W. 1988. Ecological use of vertebrate indicator species: a critique. Conservation Biology 2:316-328. [7] Gondard, H., Sandrine, J., Aronson, J. and Lavorel, S. 2003. Plant functional types: a promising tool for management and restoration of degraded lands. Applied Vegetation Science 6:223-234. [8] Kingdon, J. 1971. East African Mammals, vol. 1. Academic Press, New York. [9] Kingdon, J. 1997. The Kingdon Field Guide to African Mammals. Academic Press, London, UK. [10] Milner, J. M. 1994. Relationships between the forest dwelling people of South-West Mau and the tree hyrax, Dendrohyrax arboreus. Journal of East African Natural History, 83(1):17-29. [11] Kundaeli, J. N. 1976. Distribution of the tree hyrax (Dendrohyrax validus validus True) on Mt. Kilimanjaro, Tanzania. East African Wildlife Journal 14:253-264. [12] Nielsen, M. R. 2006. Importance, cause and effect of bushmeat hunting in the Udzungwa Mountains, Tanzania: Implications for community based wildlife management. Biological Conservation 128:509-516. [13] Gaylard, A. and Kerley, G. I. H. 2001. Habitat assessment for a rare, arboreal forest mammal, the tree hyrax Dendrohyrax arboreus. African Journal of Ecology, 39:205-212. [14] Marshall, A.R. 2007. Disturbance in the Udzungwas: Responses of Monkeys and Trees to Forest Degradation. Ph.D. Thesis, University of York, UK. [15] Marshall, A.R., Rovero, F., Struhsaker, T.T., Jørgensbye, H., Lovett, J.C. and White, P.C.L. in preparation. The species-area relationship in a threatened monkey community: controlling for externalities. American Journal of Primatology. [16] Lovett, J. C. and Pócs, T. 1993. Assessment of the conditions of the Catchment Forest Reserves, a botanical appraisal. Catchment Forestry Report 93.3. Dar es Salaam, Tanzania. [17] Topp-Jørgensen, J. E. and Pedersen, U. B. 1999. A Comparison of Mammalian Abundance in Undisturbed and Hunting Disturbed Forests of the Udzungwa Mountains, Tanzania. Unpublished M.Sc. Thesis, Zoological Museum, University of Copenhagen, Denmark. [18] Milner, J. M. and Harris, S. 1999. Habitat use and ranging behaviour of the tree hyrax, Dendrohyrax arboreus, in the Virunga Volcanoes, Uganda. African Journal of Ecology. 37(3):281-294. [19] Marshall, A.R., Aloyce, Z., Mariki, S., Jones, T., Burgess, N., Kilihama, F., Massao, J., Nashanda, E., Sawe, C., Rovero, F. and Watkin, J. 2007. Tanzania's second Nature Reserve: improving the conservation status of the Udzungwa Mountains? Oryx 41(4):429-430. [20] Dinesen, L., Lehmberg, T., Rahner, M. and Fjeldså, J. 2001. Conservation priorities for the forests of the Udzungwa Mountains, Tanzania, based on primates, duikers and birds. Biological Conservation 99:223-236. [21] Topp-Jørgensen, J. E., Brink, H. and Marshall, A. R. 2001. Large mammals in West Kilombero Scarp Forest Reserve . In: Frontier Tanzania, West Kilombero Scarp Forest Reserve - Zoological Report. Report for the Udzungwa Mountains Forest Management and Biodiversity Conservation Project, MEMA, Iringa, Tanzania, 58-68. [22] Topp-Jørgensen, J. E., Marshall, A. R. and Brink, H. 2001. Assessment of the impact of human forest use in New Dabaga/ Ulangambi Forest Reserve. In: Frontier Tanzania, New Dabaga/Ulangambi Forest Reserve - Botanical and Forest Use Report. Report for the Udzungwa Mountains Forest Management and Biodiversity Conservation Project, MEMA, Iringa, Tanzania . [23] Ramsey, F. L. and Scott, J. M. 1979. Estimating population densities from variable circular plot surveys. Statistical Ecology Series 5:155-181. [24] Reynolds, C. M., Scott, J. M. and Nussbaum, R. A. 1980. A variable circular plot method for estimating bird numbers. Condor 82:309-313. [25] Marshall, A.R., Lovett, J.C. and White, P.C.L. 2008. Selection of line-transect methods for estimating the density of group-living animals: lessons from the primates. American Journal of Primatology 70:1-11. [26] Milner, J. M. and Harris, S. 1999. Activity patterns and feeding behaviour of the tree hyrax, Dendrohyrax arboreus, in the Parc National des Volcans, Rwanda. African Journal of Ecology 37(3):267-280. [27] Estes, R. D. 1991. The Behaviour Guide to African Mammals. University of California Press, Berkely and Los Angeles, California, USA. pp.xxii+611. [28] Rovero, F. & Marshall, A.R. 2004. Estimating the abundance of forest antelopes by using line transect techniques: a case from the Udzungwa Mountains of Tanzania. Tropical Zoology 17:267-277. [29] Bowkett, A., Rovero, F., Marshall, A.R. 2007. The use of camera-trap data to model habitat selection by antelope species in the Udzungwa Mountain forests, Tanzania. African Journal of Ecology . On-line doi: 10.1111/j.1365-2028.2007.00881.x [30] Richard, P. B. 1964. Notes sur la cri du Dendrohyrax dorsalis (Hyracoidea). Biol.Gabon. 1:73-84. [31] Jones, C. 1978. Dendrohyrax dorsalis. Mammalian Species 113:1-4. [32] Rudnai, J. 1984. Activity cycle and space utilisation in captive Dendrohyrax arboreus. South African Journal of Zoology 19:124-128. [33] Marshall, A.R., Topp-Jørgensen, J.E., Brink, H., Fanning, E. 2005. Monkey abundance and social structure in two high elevation forest reserves in the Udzungwa Mountains of Tanzania. International Journal of Primatology 26:127-145. [34] Buckland, S.T., Anderson, D.R., Burnham, K.P., Laake, J.L. 1993. Distance Sampling: Estimating Abundance of Biological Populations. London: Chapman & Hall. [35] Buckland, S.T., Anderson, D.R., Burnham, K.P., Laake, J.L., Borchers, D.L., Thomas, L. 2001. Introduction to Distance Sampling: Estimating Abundance of Biological Populations. Oxford: Oxford University Press. [36] Burnham, K.P., Anderson, D.R., Laake, J.L. 1980. Estimation of density from line-transect sampling of biological populations. Wildlife Monographs 72, Supplement to the Journal of Wildlife Management 44. [37] Burgess, N.D., Loucks, C., Stolton, S., Dudley, N. 2007. The potential of forest reserves for augmenting the protected area network in Africa. Oryx 41:151-159. Comments Reader comments are generally moderated. If you find something inappropriate, please contact Tropical Conservation Science. The opinions expressed in reader comments are those of the author only, and do not necessarily reflect the opinions of other authors or Tropical Conservation Science. |
 |
Tropical Conservation Science is an open-access e-journal that publishes research relating to conservation of tropical forests and other tropical ecosystems.
Volume 1: Issue 1 Table of Contents Articles PDF version General interest review All issues Mar 2008 Jun 2008 Sep 2008 Dec 2008 Mar 2009 Jun 2009 Sep 2009 Dec 2009 Mar 2010 Jun 2010 Sep 2010 Dec 2010 Mar 2011 Jun 2011 Sep 2011 Dec 2011 Mar 2012 Jun 2012 Sep 2012 Dec 2012 Mar 2013 Jun 2013 Aug 2013 Sep 2013 Nov 2013 Dec 2013 Mar 2014 Jun 2014 Sep 2014 Dec 2014 Mar 2015 Jun 2015 Sep 2015 Dec 2015 Mar 2016 Jun 2016 Most downloaded 2008 2009 2010 2011 2012 All time ADVERTISEMENT SEARCH  This work is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||


|
About | Privacy Copyright mongabay.com 2008-2014 |